
Over the next while I plan to blog about various papers that have come out of our research program. I won’t get to all of them, obviously. But I do plan to pick and choose a few recent ones, and/or ones that have been highlights to this point in my career.
I’m going to begin with a very recent paper from my lab on bark beetle larval overwintering physiology. The paper is entitled “Global and comparative proteomic profiling of overwintering and developing mountain pine beetle, Dendroctonus ponderosae (Coleoptera: Curculionidae), larvae” and is available in open access here.
Context: Mountain pine beetles usually spend their winters as small, young larvae under the bark of their host tree. In this location, they are exposed to extremely cold temperatures, sometimes ranging below –30°C and even pushing down towards -40°C. Mountain pine beetle larvae survive those temperatures by resisting freezing. Sometime in the autumn they begin to accumulate at least one antifreeze compound (glycerol) in their bodies, and then in the spring they presumably return that antifreeze compound (and perhaps others) to general metabolism for energy to complete their development
Cold temperatures have historically limited the range of the mountain pine beetle both in terms of longitude and latitude, and in terms of elevation. However, climate change has reduced the probability of cold winter temperatures – particularly the probability of extreme cold events fairly early in the autumn or fairly late in the spring. At those ‘shoulder seasons’ the larval insects have either not accumulated enough antifreeze compounds in their tissues (autumn, around Hallowe’en) or have metabolized most of it (spring, around Easter). Those are the vulnerable periods, and deep cold at those can cause populations to crash rapidly.
The lack of unseasonal cold events or of generally very deep cold in the heart of the winter over the past years has been one factor that has driven the dramatic outbreak that we’ve seen in British Columbia. In addition, historically colder areas such as the eastern slopes of the Rockies and central Alberta or high elevation areas in the Rockies have not been as cold either. This has allowed mountain pine beetles to survive winters and to move into hosts, such as jack pine and whitebark pine, that they have not historically used in the recorded past. In the case of jack pine outbreaks, the fear is that the beetle, freed from its main confine on west slope of the Rockies, is poised to move across Canada’s boreal forest. In the case of whitebark pine, the insect may further endanger already-threatened trees that are important to higher alpine ecosystems.
What we did: Up until now, the main known antifreeze compound in mountain pine beetle larva has been glycerol. We suspected that there was more to the insect’s overwintering physiology than just that, as most insects use several strategies to avoid freezing. So we conducted a proteomics experiment. That means that we surveyed the levels of all of the proteins in early-autumn larvae and compared them to levels of proteins in late-autumn larvae to look for changes. Similarly, we compared the levels of all detectable proteins between early-spring and late-spring larvae. Because we now have copious amounts of genomic data for the mountain pine beetle, we could identify which proteins did what in the insect and we could draw some conclusions as to which metabolic pathways and physiological processes were activated or deactivated in overwintering larvae at different times of the year.
What we found: In total we found 1507 proteins in all of our larval samples. Of these, 33 either increased or decreased in their levels between early- and late-autumn and 473 either increased or decreased in their levels between early- and late spring. Of the proteins that were present in either increased or decreased levels in one of the two seasons, 18 of them showed such changes in both seasons. This Venn diagram from the paper shows this general result:
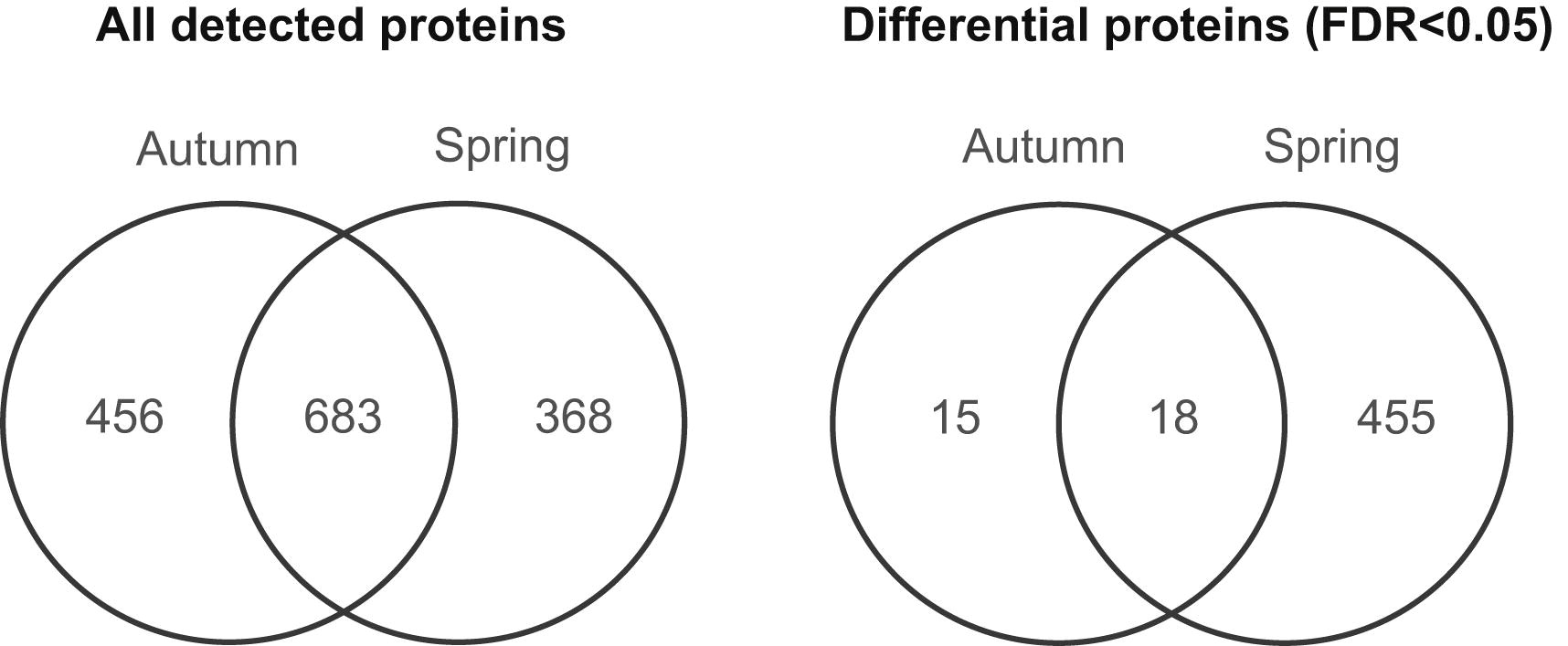
These proteins can be classified into a number of general functional groups, as seen in this pie chart from the paper:
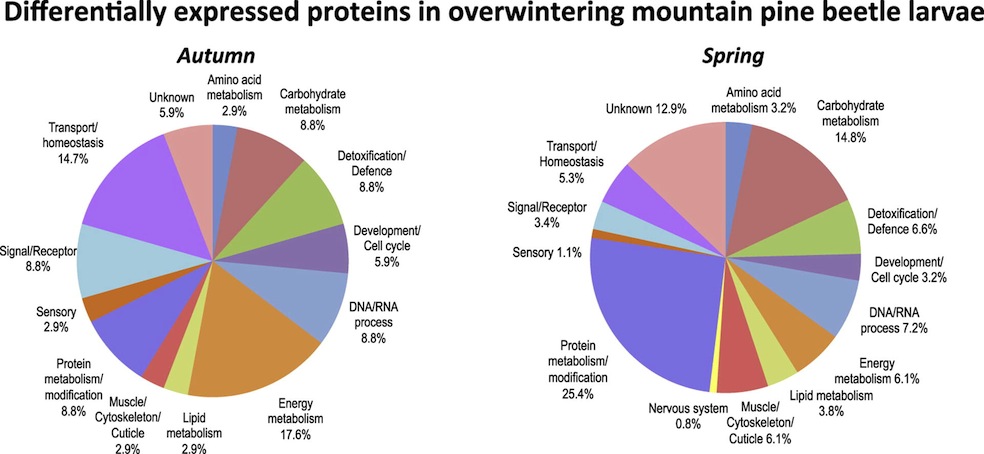
Of course, large groupings are not as informative as looking at individual proteins. So that is what we did, as I will write about in the next section.
What this means: In proteomics work like this, when we are dealing with hundreds of proteins, it is obvious that there is so much complexity that it would take untold pixels to explain everything. In fact, like may ‘-omics’ studies, the original authors (us, in this case) have to pick and choose things that seem interesting to them and then leave it to others wearing different research glasses to find other interesting trends. What follows are a few highlights that we noticed in the context of our research program. Our hope is that others will take our data and find other interesting things that we may have missed.
Glycerol: Our results confirm past work implicating glycerol as an important antifreeze compound in the mountain pine beetle. The data also confirm previous work in our lab (Fraser 2011, referenced in the paper) that shows certain glycerol biosynthetic genes being upregulated in the autumn and downregulated in the spring. Of particular note were the extreme variations in an enzyme called PEPCK (phosphoenolpyruvate carboxykinase) which likely indicates some level of nutritional stress in larvae heading into the cold of winter.
Trehalose: Trehalose is a major hemolymph (insect “blood”) sugar, and it has been found to be important in insect cold tolerance in other species. The levels of an enzyme involved in trehalose biosynthesis increased significantly in the autumn and decreased significantly in the spring, indicating that trehalose might function alongside glycerol as an antifreeze compound.
2-deoxyglucose: The largest autumn increases and spring decreases for any protein that we observed was for one enzyme that is involved in the biosynthesis of 2-deoxyglucose. By looking at what 2-deoxyglucose does in other organisms, we can make some guesses as to what it is doing in the mountain pine beetle. It is possible that 2-deoxyglucose regulates larval metabolism to direct energy flow appropriately toward overwintering in the autumn; that it acts in stress physiology as the insect enters a difficult period of its life; or that it is functional as an antifreeze compound. It’s also possible that it functions in more than of these roles. What is clear is that this metabolite, not previously detected in this species, is likely very important in mountain pine beetle overwintering physiology. So we have some work on our hands to figure out exactly what it’s doing.
Stress, in general: The levels of a number of proteins associated with stress physiology – for instance ferritin, superoxide dismutase and phospholipid hydroperoxide glutathione peroxidase – increased in the autumn and, in some cases, decreased again in the spring. The fact that winter is a stressful period in a mountain pine beetle’s life cycle is obvious from the basic ecology of the organism. We now have a number of stress physiology protein targets to investigate in further research.
Energy use during development: The increases and decreases of particular enzymes involved in basic metabolism indicate that mountain pine beetle larvae put most of their resources into overwintering preparation in the autumn, and only when they have survived to the spring do they begin to divert resources to ongoing developmental processes.
Detoxification of host defenses: A number of proteins commonly involved in detoxification of host chemical defenses were present in autumn larvae but, for the most part, showed reductions in the larvae as the spring progressed. Previous work in our lab has shown that larvae in the late-summer experience extremely high levels of host defense compounds. So autumn larvae are working hard to get prepared for overwintering while also dealing with a toxic environment. Once the winter is over, and the host tree is long dead, it is likely that residual host toxins have either been removed by the beetle’s symbiotic fungi or that they have naturally degraded or dissipated. In any case, the detoxification enzymes are seeming not needed to nearly the degree in the late spring that they were during the autumn. The larvae that survive living in a toxic wasteland in the autumn and that do not freeze to death in the winter are then free to use remaining stores of energy plus whatever they can glean from their host tree to complete their developmental cycle through the spring and early-summer.
Why this is important: This is the first comprehensive look at what is going on in an overwintering bark beetle. While there has been a bit of previous physiological work on mountain pine beetles and a few other bark beetle species, our work in the Tria Project has moved us into the post-genomic era for the mountain pine beetle. That means that we have an extensive genomic database and that we can conduct experiments like this that reveal the workings of a number of physiological systems all at once. We are doing other ‘-omics’ work as well on overwintering mountain pine beetle larvae, including transcriptomics (monitoring messenger RNA levels during different seasons) and directed metabolomics (monitoring specific metabolites related to overwintering) work. And we are doing experiments where we track the expression of specific genes and the activity of specific enzymes revealed to be important during this phase of the insect’s life cycle. Of course our lab, alone, can’t do all of the experimentation suggested by these results. In fact, the data are so extensive that we can’t even conceive of all of the potential experiments. That is what is cool about ‘-omics’ research – there’s no telling who will look at it and think “ah ha! I have a great idea!”
Ultimately we hope that this paper has blown the door open on bark beetle overwintering physiology. Further research is bound to uncover new and interesting results, and since winter cold and climate change play such a large role in the growth of mountain pine beetle populations, such results will help us to understand better where and how the beetles are spreading into new regions and new, susceptible hosts.
Where we are going with this: As I mentioned above, the amount of data from this one study is staggering. This is our lab’s first publication from the larger Tria Project and there are others in the works. Some of them will also produce similar copious data. Others have been designed to look at specific small portions of this study and of some of our other data. We are currently focusing in on some of the metabolic pathways and physiological processes that I mentioned above. And we hope that others are able to take our data and use it for different analyses. For instance, we have surveyed protein levels across much of the larval developmental period. Perhaps others interested in insect development will find and be able to use new information on development in the Coleoptera (beetles) generally, and in bark beetles and other weevils specifically.
This was a really fun study. We certainly hope that the data will be as useful to others as it has been for us already. This work has also moved our research program firmly into the realm of insect overwintering research, and it has been a great introduction for us into proteomics and the era of “big data” in the biological sciences.
![]()
Bonnett TR, Robert JA, Pitt C, Fraser JD, Keeling CI, Bohlmann J, & Huber DP (2012). Global and comparative proteomic profiling of overwintering and developing mountain pine beetle, Dendroctonus ponderosae (Coleoptera: Curculionidae), larvae. Insect biochemistry and molecular biology, 42 (12), 890-901 PMID: 22982448